ΣΥΛΛΟΓΟΣ (OR "ΣΥΝΔΕΣΗ" "" ΣΥΝΔΕΣΜΟΣ ")
Μέχρι τώρα αντιμετωπίζαμε τη διασταύρωση Mendelian dihybrid (ή polyhybrid) υποθέτοντας ότι τα διαφορετικά αλληλόμορφα ζευγάρια βρίσκονται στην πραγματικότητα σε διαφορετικά ομόλογα ζεύγη χρωμοσωμάτων. Αλλά ο αριθμός των χρωμοσωμικών ζευγών, αν και διαφορετικός από είδος σε είδος, ποικίλλει σε στενά όρια φτάνουν περίπου τα εκατό χρωμοσώματα), ενώ ο αριθμός των γονιδίων μπορεί να μετρηθεί σε δεκάδες χιλιάδες.
Ότι οι χαρακτήρες που επέλεξε ο Mendel για τα πειράματά του διαχωρίστηκαν ανεξάρτητα (χωρίς να συγχέουμε τους υπολογισμούς στην κατανομή των φαινοτύπων στο F2 του πολυυβριδικού σταυρού) ήταν μια τυχερή ευκαιρία. Εάν δύο ζεύγη αλληλόμορφων βρέθηκαν σε γειτονικούς τόπους, ο νόμος θα ακολουθούσε θα ονομαζόταν νόμος της ένωσης.
Γνωρίζοντας ότι πολλοί χαρακτήρες έχουν τον τόπο τους σε ένα μόνο χρωμόσωμα και ότι είναι τα χρωμοσωμικά ζεύγη που διαχωρίζονται ανεξάρτητα στη μείωση, μπορεί να γίνει κατανοητό πόσο συχνά συμβαίνει δύο ζεύγη χαρακτήρων, εάν σχετίζονται με ένα χρωμόσωμα στο γονικό οργανισμός, παραμένουν συνδεδεμένοι εξίσου και στον γαμέτη και επομένως στον οργανισμό στον οποίο θα φέρει το δικό του γενετικό υλικό.
Έτσι, βλέπουμε ότι η "ένωση αντιπροσωπεύει" μια εξαίρεση, κάθε άλλο παρά σπάνια, στην ανεξαρτησία που αναφέρεται στον τρίτο νόμο του Mendel.
ΑΝΤΑΛΛΑΓΗ OR "ΠΕΡΙΣΣΟΤΕΡΑ" ΚΑΙ ΑΝΑΔΟΣΗ
Μιλώντας για μείωση έχουμε δείξει ότι υπάρχουν δύο διαφορετικές στιγμές ανάμειξης του γενετικού υλικού: η μία είναι αυτή του διαχωρισμού των χρωμοσωμάτων στους γαμέτες και αυτή είναι που παρατηρήθηκε από τον Mendel.
Η άλλη στιγμή, που πραγματικά προηγείται, είναι εκείνη στην οποία οι τέσσερις χρωματίδες κάθε ζεύγους ομόλογων χρωμοσωμάτων ανταλλάσσουν αμοιβαία ταυτόσημα χαρακτηριστικά. Μετά από αυτήν την ανταλλαγή, δύο παράγοντες που σχετίζονται με το ίδιο χρωμόσωμα θα είναι αντίθετα ανεξάρτητοι στους γαμέτες. ότι μια ανταλλαγή λαμβάνει χώρα είναι ανάλογη, με μια πρώτη προσέγγιση, με το μήκος του χρωμοσώματος, και σε μακρύτερα χρωμοσώματα μπορεί να υπάρξει ακόμη περισσότερο από, μια ανταλλαγή.
Το φαινόμενο μπορεί να ανιχνευθεί κυτταρολογικά, παρατηρώντας αρκετό αριθμό μειωμάτων κάτω από το μικροσκόπιο.
Ο ρυθμός επανάληψης είναι ο ρυθμός με τον οποίο οι δύο χαρακτήρες που σχετίζονται με τη γονική γενιά ανασυνδυάζονται διαφορετικά στο F2.
Εάν οι δύο τόποι είναι απολύτως συνεχόμενοι, η πιθανότητα να τους χωρίσει ένας χιασμός θα είναι πρακτικά μηδενική. Ο ρυθμός ανασυνδυασμού θα είναι: n ° ανασυνδυασμένοι. Εάν δύο τόποι βρίσκονται σε δύο διαφορετικά χρωμοσώματα, το ποσοστό ανασυνδυασμού θα είναι 0,5 (ίση πιθανότητα, για δύο χαρακτήρες που ενώθηκαν στη γενιά Ρ, να βρεθούν τυχαία μαζί στο F2). Ο ρυθμός ανασυνδυασμού μπορεί επομένως να κυμαίνεται μεταξύ 0,0 και 0,5. Για μικρές αποστάσεις στο χρωμόσωμα, η απόσταση και ο ρυθμός ανασυνδυασμού είναι ευθέως ανάλογοι. Για μεγαλύτερες αποστάσεις υπάρχει η πιθανότητα να πραγματοποιηθούν δύο ανταλλαγές μεταξύ δύο τόπων. Τώρα θα φανεί σαφές ότι δύο παράγοντες που χωρίζονται από δύο ανταλλαγές συνδέονται ξανά. είναι σαφές, σε αυτό το σημείο, ότι η αναλογικότητα μεταξύ της απόστασης των τόπων και της πιθανότητας ανασυνδυασμού χάνεται.
Οι εστίες που βρέθηκαν συνδεδεμένες στο ίδιο χρωμόσωμα αποτελούν «ομάδες συσχέτισης». Οι πολύ μακρινοί τόποι μπορεί να έχουν τέτοια πιθανότητα διαχωρισμού με ανταλλαγή ώστε να συμπεριφέρονται ως ανεξάρτητοι, αλλά καθένας από αυτούς θα σχετίζεται, με χαμηλότερο ποσοστό ανασυνδυασμού, με τους ενδιάμεσους τόπους.
Όταν είναι γνωστά τα ποσοστά ανασυνδυασμού μεταξύ πολλών ζευγών γονιδίων σε μια ομάδα συσχέτισης, μπορεί να ξεκινήσει η κατασκευή «γενετικών χαρτών». Έχοντας υπόψη ότι η απόσταση μεταξύ δύο γονιδίων (a και b) εκφράζεται με το ρυθμό ανασυνδυασμού και ότι η απόσταση a από ένα τρίτο γονίδιο c μπορεί να είναι είτε το άθροισμα είτε η διαφορά ως προς την απόσταση του από το b, είναι πιθανό για την ανασυγκρότηση ενός χάρτη των αμοιβαίων αποστάσεων, ο οποίος θα είναι ο γενετικός χάρτης εντός αυτής της ομάδας συσχέτισης, δηλαδή αυτού του χρωμοσώματος.
Πρέπει τώρα να εξετάσουμε γενικά ορισμένες έννοιες που περιορίζουν τη φαινοτυπική εκδήλωση των γονότυπων χαρακτήρων.
Πρώτα απ 'όλα θα μιλήσουμε για τις έννοιες της διείσδυσης και της εκφραστικότητας και στη συνέχεια θα αφιερώσουμε ιδιαίτερη προσοχή στα φαινόμενα ρύθμισης της γονιδιακής δράσης.
ΔΙΑΔΟΣΗ
Η διείσδυση ενός γονιδίου αντιπροσωπεύει την ικανότητά του να εκδηλώνεται στον φαινότυπο. Η διείσδυση μετριέται στατιστικά μετρώντας τη συχνότητα των φαινοτύπων που δείχνουν αυτόν τον χαρακτήρα από 100 γονότυπους που τον περιέχουν. Ένα χαρακτηριστικό με διείσδυση 0,7 είναι ένα χαρακτηριστικό που εμφανίζεται φαινοτυπικά στο 70% της γονότυπης συχνότητάς του.
ΕΚΦΡΑΣΗ
Η εκφραστικότητα είναι μια ποσοτική εκτίμηση του βαθμού φαινοτυπικής εκδήλωσης.
ΚΑΝΟΝΙΣΜΟΣ ΓΕΝΙΚΗΣ ΔΡΑΣΗΣ
Τα κύτταρα παράγουν όλα τους τα ένζυμα και τις πρωτεΐνες τους με την ίδια ταχύτητα και ταυτόχρονα. Τα κύτταρα Escherichia coli, για παράδειγμα, μπορούν να τροφοδοτηθούν με ενέργεια και άτομα άνθρακα από τον δισακχαρίτη λακτόζης καθώς είναι σε θέση να τα διασπάσουν σε γλυκόζη και γαλακτόζη χάρη στο ένζυμο βήτα-γαλακτοσιδάση. Σε ένα φυσιολογικό E. coli που μπορεί να έχει λακτόζη, υπάρχουν περίπου 3.000 μόρια βήτα-γαλακτοσιδάσης, ίσα με το 3% των πρωτεϊνών αυτού του κυττάρου. απουσία λακτόζης θα υπάρχει μόνο ένα μόριο βήτα-γαλακτοσιδάσης ανά βακτηριακό κύτταρο. Η γαλακτοσιδάση θα συντεθεί από νέα μόρια mRNA όταν μπορεί να χρησιμοποιηθεί. Μεταλλαγμένα στελέχη του E. coli πλούσια σε ένζυμο είναι γνωστά ακόμη και όταν απουσιάζει η λακτόζη: αυτά τα μεταλλαγμένα μειονεκτούν σε σύγκριση με τα κανονικά κύτταρα καθώς αναγκάζονται σε άσκοπη κατανάλωση ενέργειας και υλικών για την παραγωγή του ενζύμου το οποίο θα παραμείνει χωρίς υπόστρωμα. Ουσίες που προκαλούν αύξηση της ποσότητας του ενζύμου, όπως συμβαίνει με τη λακτόζη, θα ονομάζονται επαγωγείς, ενώ τα ένζυμα θα λέγονται ότι είναι επαγώγιμα. Άλλες ουσίες προκαλούν, επίσης αυτές με έναν συγκεκριμένο τρόπο, την παραγωγή ορισμένων ενζύμων. Επίσης, στο E. coli, για παράδειγμα, ικανό να κατασκευάσει όλα τα αμινοξέα του, έχοντας άνθρακα και αμμώνιο (NH3), η παρουσία στο μέσο καλλιέργειας ενός συγκεκριμένου αμινοξέος (ιστιδίνη, για παράδειγμα) εμποδίζει την παραγωγή όλων των ενζύμων που σχετίζονται με η βιοσύνθεση του ίδιου του αμινοξέος: θα πούμε για αυτά τα ένζυμα ότι είναι κατασταλτικά. Στα βακτηριακά κύτταρα τα μόρια του mRNA κατεδαφίζονται λίγο μετά τον σχηματισμό τους και γι 'αυτό ο έλεγχος της παραγωγής του mRNA σημαίνει τον έλεγχο της ενζυματικής σύνθεσης ταυτόχρονα ώρα
ΤΟ ΟΠΕΡΟΝ
Για να εξηγήσει πώς το βακτηριακό κύτταρο είναι σε θέση να ελέγξει τη δική του παραγωγή ενζύμων, ο Jacob και ο Monod διατύπωσαν την υπόθεση του οπερονίου. Το οπερόνιο σχηματίζεται από πολλά γονίδια που είναι λειτουργικά συγγενικά και ευθυγραμμισμένα χωρίς ασυνέχεια κατά μήκος ενός τμήματος DNA. Το οπερόνιο αποτελείται από τρεις διαφορετικούς τύπους γονιδίων: τον προαγωγό, όπου αρχίζει ο σχηματισμός του mRNA ο χειριστής, όπου ασκείται ο έλεγχος · ένα ή περισσότερα δομικά γονίδια, τα οποία κωδικοποιούν ένζυμα ή άλλες πρωτεΐνες. Στο σύστημα της β-γαλακτοσιδάσης, το οπερόνιο περιλαμβάνει, εκτός από αυτό για τη βήτα-γαλακτοσιδάση, επίσης δύο άλλα γονίδια δομικών κωδικοποιήσεων για άλλα ένζυμα που εμπλέκονται στο μεταβολισμό της λακτόζης.Αυτά τα γονίδια είναι το ένα δίπλα στο άλλο και μεταγράφονται το ένα μετά το άλλο κατά μήκος της ίδιας έλικας DNA σε ένα μόνο μόριο mRNA. Τα μόρια mRNA που παράγονται έτσι είναι ενεργά για πολύ σύντομο χρονικό διάστημα, μετά το οποίο καταστρέφονται από συγκεκριμένα ένζυμα.
Η δραστηριότητα του οπερονίου με τη σειρά του ελέγχεται από ένα άλλο γονίδιο, τον ρυθμιστή, το οποίο μπορεί επίσης να απέχει από το οπερόνιο: αυτός ο ρυθμιστής κωδικοποιεί μια πρωτεΐνη, που ονομάζεται καταστολέας, η οποία φαίνεται να συνδέεται με το DNA στο γονίδιο χειριστή. μεταξύ του προαγωγέα και των δομικών γονιδίων εμποδίζει την παραγωγή mRNA.
Ο καταστολέας με τη σειρά του ελέγχεται και ο έλεγχος πραγματοποιείται μέσω μιας ουσίας "σήματος". Στην περίπτωση επαγώγιμων ενζύμων, αυτή η ουσία είναι ο "επαγωγέας. Ο επαγωγέας" συνδέεται με το μόριο καταστολής τροποποιώντας το σχήμα του, έτσι ώστε να μην μπορεί πλέον να προσαρμοστεί στο DNA: στην περίπτωση αυτή, αφού δεν υπάρχει καταστολέας μεταξύ του προαγωγού και των δομικών γονιδίων , ο καταστολέας μπορεί να σχηματίσει τα μόρια mRNA και από αυτά τα μόρια πρωτεΐνης. Με την εξάντληση της παροχής του επαγωγέα, ο ρυθμιστής θα ανακτήσει ξανά τον έλεγχο, ο οποίος θα σταματήσει την παραγωγή νέου mRNA, επομένως νέων πρωτεϊνών. Στο σύστημα της β-γαλακτοσιδάσης ο επαγωγέας είναι η λακτόζη ή μια ουσία πολύ παρόμοια με αυτήν. θα ενωθεί με τον καταστολέα απενεργοποιώντας το έτσι ώστε να επιτρέψει τη βιοσύνθεση ενζύμων. Στην περίπτωση των κατασταλτικών ενζύμων, η ουσία που δρα ως «σήμα» δρα ως πυρήνας: ο καταστολέας είναι ενεργός μόνο εάν συνδυάζεται με τον πυρήνα πυρήνα. Στο σύστημα ιστιδίνης, το οποίο περιλαμβάνει δώδεκα διαφορετικά ένζυμα, είναι αυτό το αμινοξύ, σε συνδυασμό με το tRNA, τον πυρήνα πυρήνα ιστιδίνη.
ΑΛΟΣΤΕΡΙΚΕΣ ΑΛΛΗΛΕΠΙΔΡΑΣΕΙΣ
Οι αλλοστερικές αλληλεπιδράσεις, που περιλαμβάνουν την απενεργοποίηση ενός ενζύμου αλλάζοντας το σχήμα του, παρέχουν έναν διαφορετικό τρόπο ρύθμισης της μεταβολικής δραστηριότητας ενός κυττάρου. Οι αλλοστερικές αλληλεπιδράσεις επιτρέπουν τον ακριβέστερο έλεγχο από το σύστημα επαγωγέα -καταστολέα του οπερονίου, αλλά δεν επιτυγχάνουν το χρήσιμο αποτέλεσμα του αποκλεισμού της βιοσύνθεσης μιας δεδομένης ουσίας από το πρώτο στάδιο - την παραγωγή ενός mRNA.
ΣΥΣΤΗΜΑΤΑ ΕΛΕΓΧΟΥ ΣΤΟ EUCARIUS
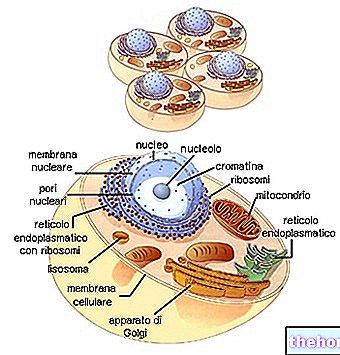
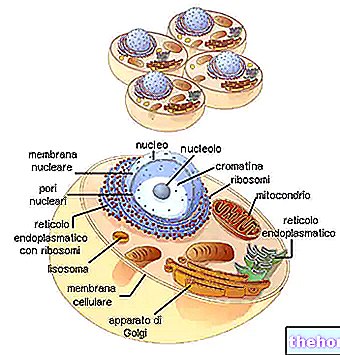
Υπάρχουν ορισμένα γεγονότα που οδηγούν στο συμπέρασμα ότι ένα σύστημα ρύθμισης παρόμοιο με το οπερόνιο λειτουργεί και κυριαρχεί μεταξύ των φυτών και των ζώων. Τα χρωμοσώματα αυτών των οργανισμών διαφέρουν σε μεγάλο βαθμό από αυτά του E. coli και άλλων προκαρυωτικών. Ο έλεγχος των γονιδίων σε αυτά Ο μηχανισμός μίτωσης είναι τέτοιος ώστε κάθε κύτταρο ενός δεδομένου φυτού ή ζώου να διαθέτει όλες τις πληροφορίες
γενετικά που υπάρχουν στο γονιμοποιημένο ωάριο. Επομένως, τα περισσότερα γονίδια σε οποιοδήποτε εξειδικευμένο κύτταρο θα παραμείνουν αναποτελεσματικά καθ 'όλη τη διάρκεια ζωής του κυττάρου. Το DNA σε αυτά τα κύτταρα συνδέεται πάντα με πρωτεΐνες. Επομένως, είναι πιθανό ότι η γονιδιακή καταστολή στους ευκαρυωτικούς οργανισμούς απαιτεί ακριβώς αυτόν τον συσχετισμό μεταξύ DNA και πρωτεϊνών.












.jpg)